![]()
![]()
![]()
![]()
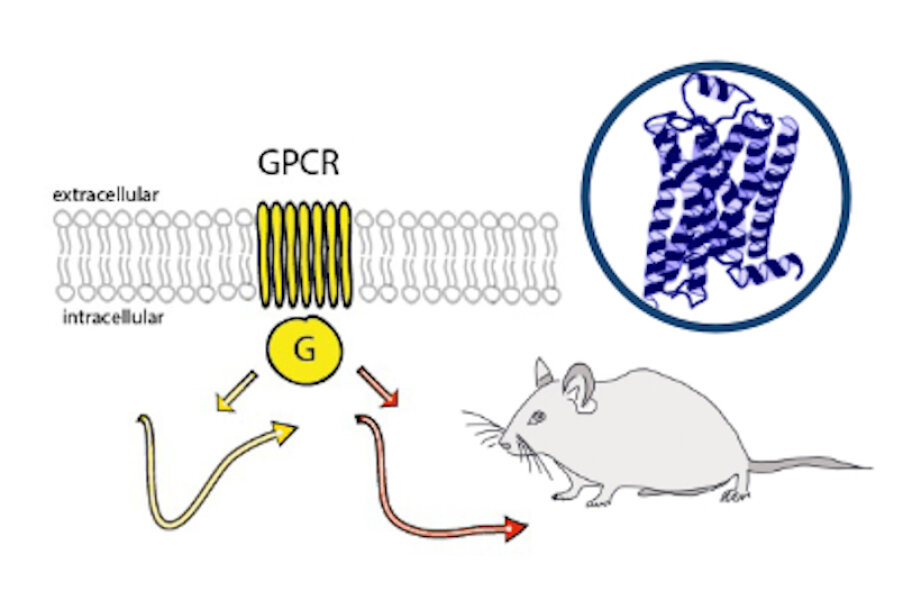
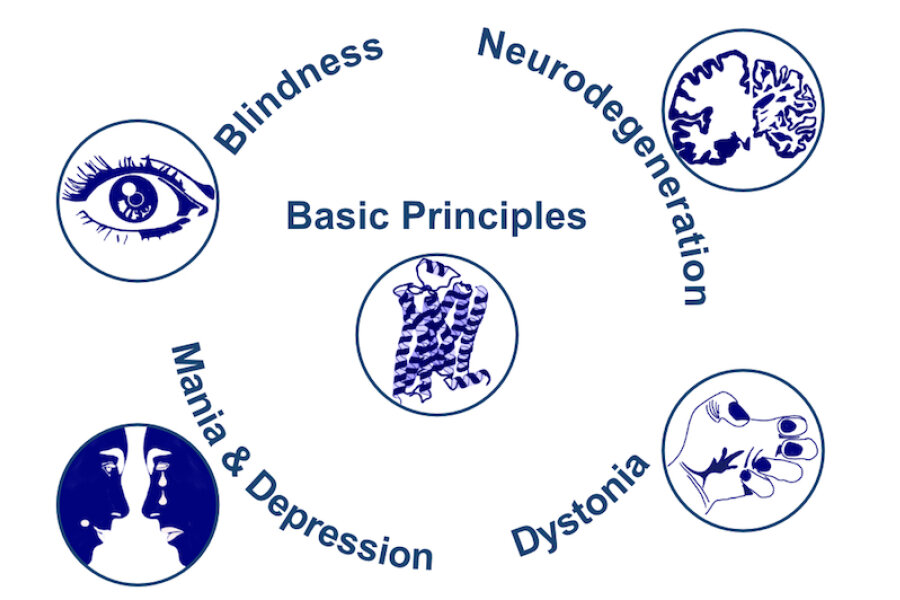
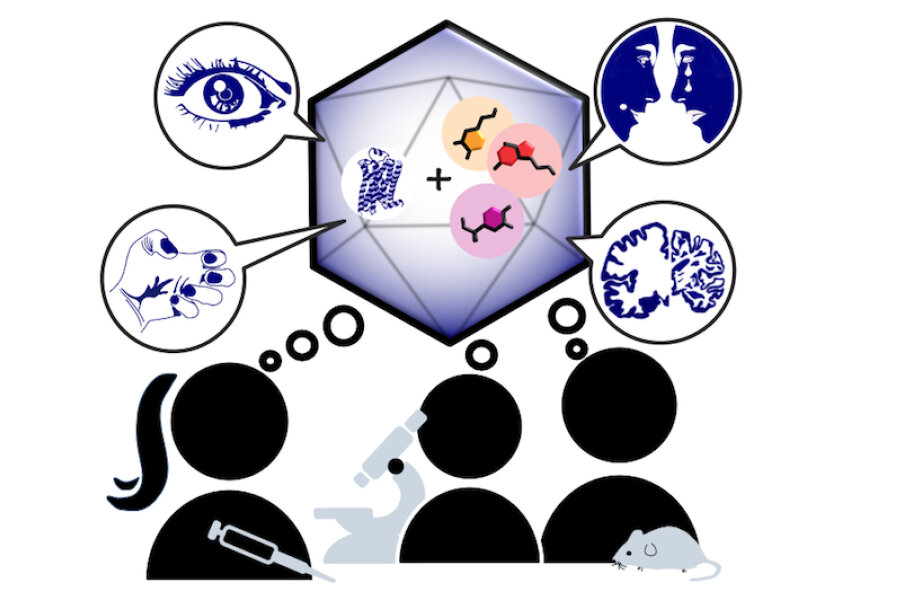
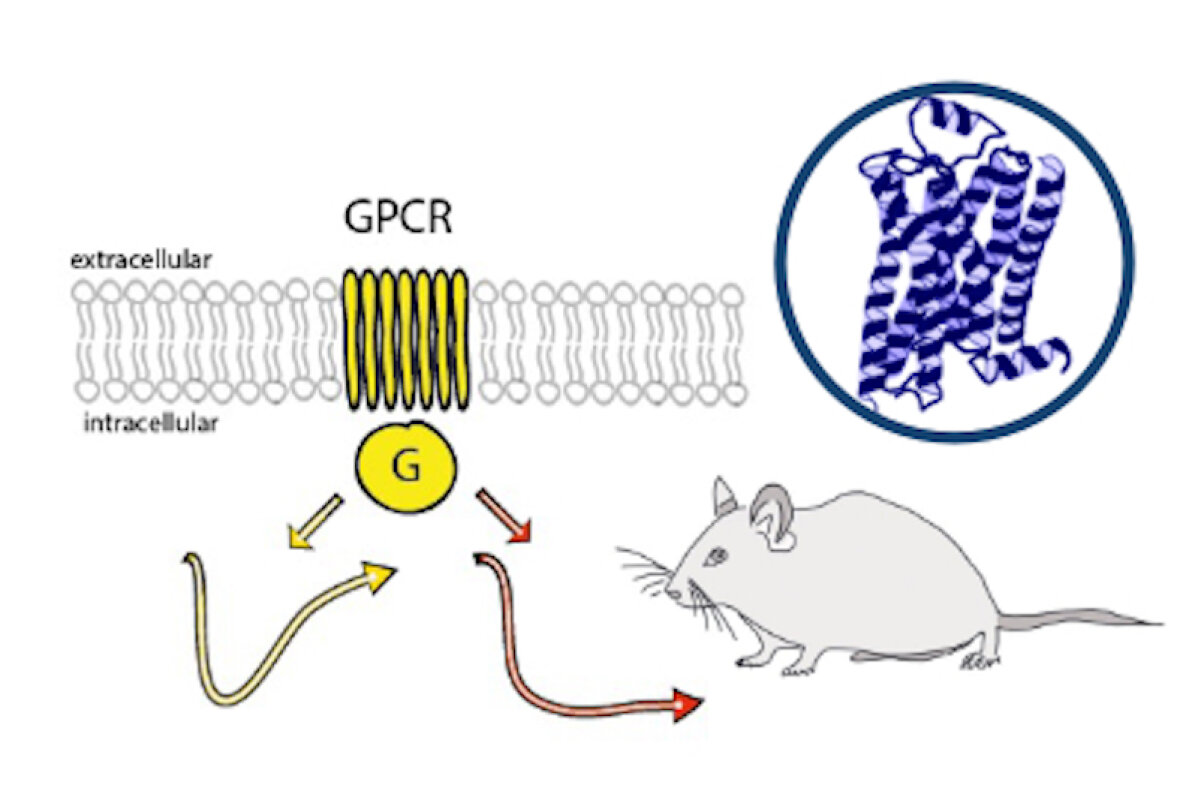
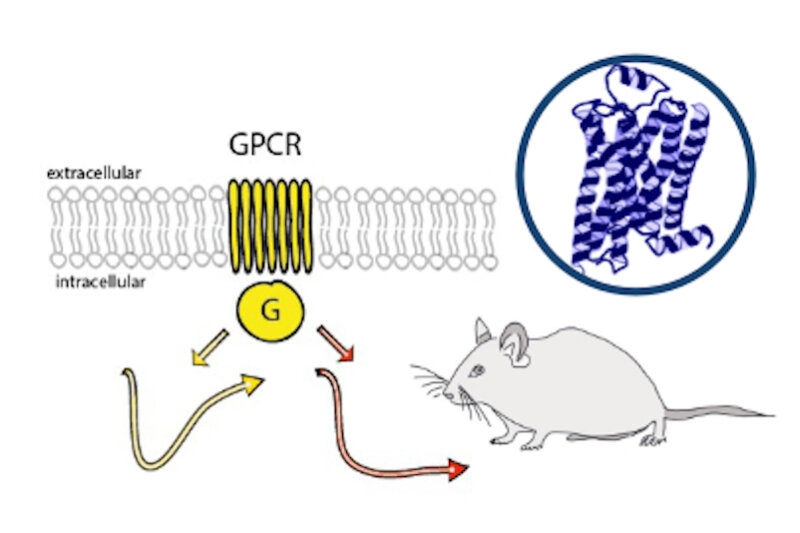
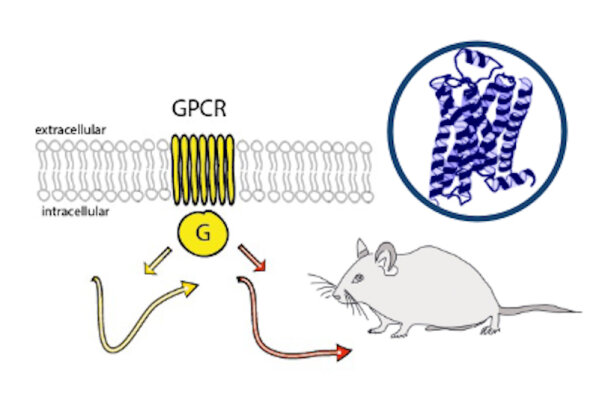
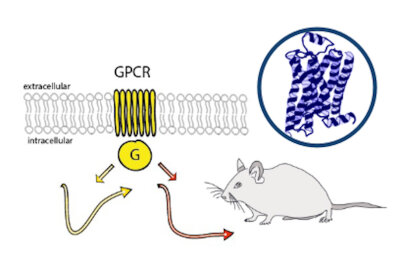
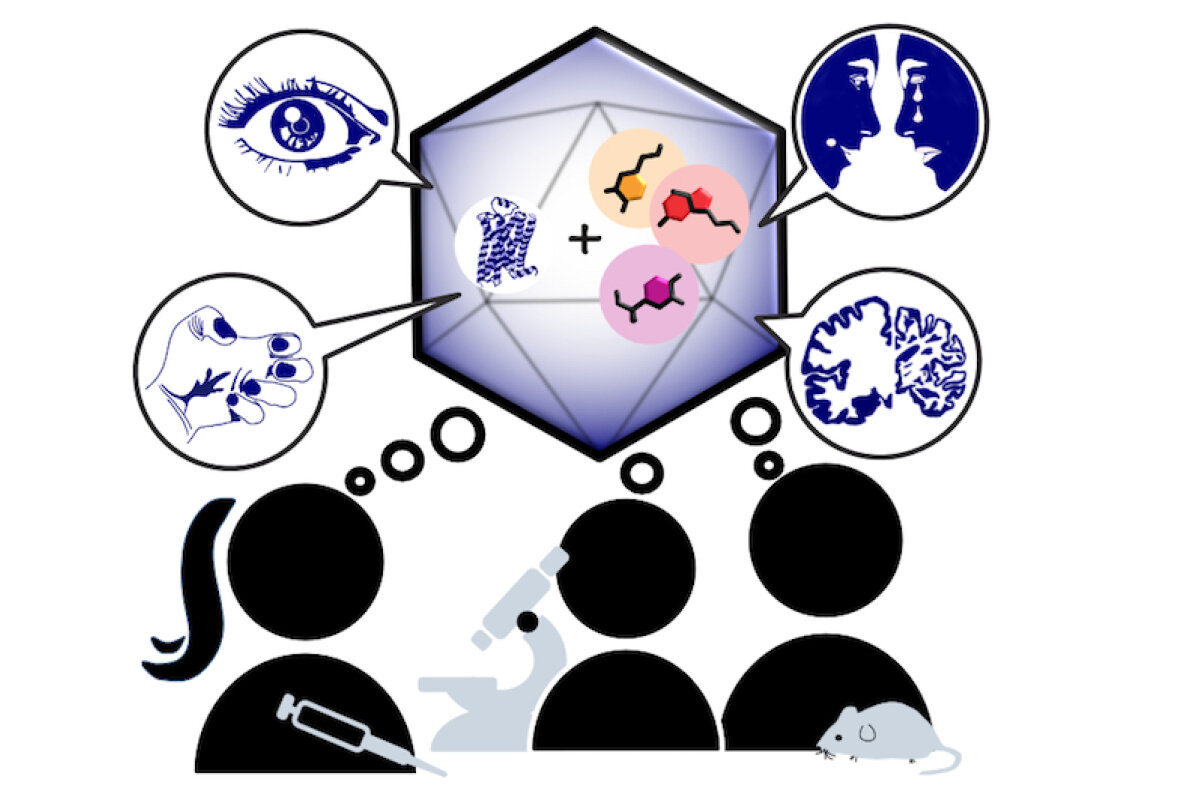
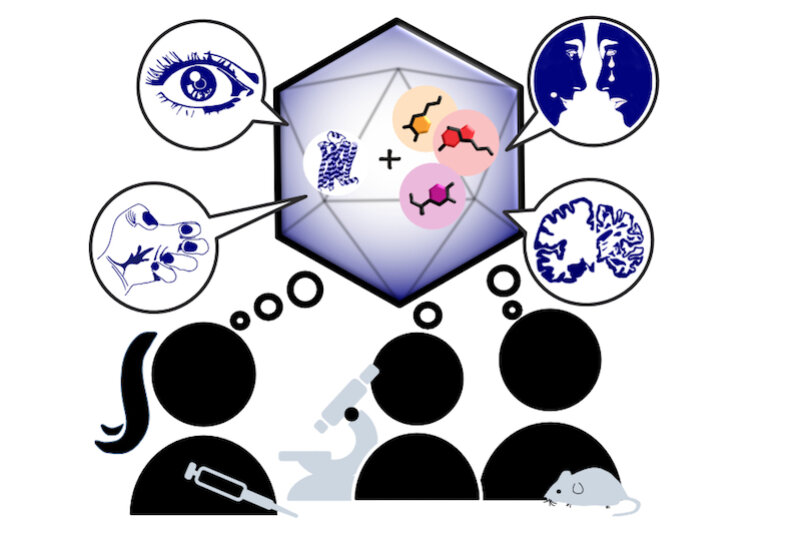
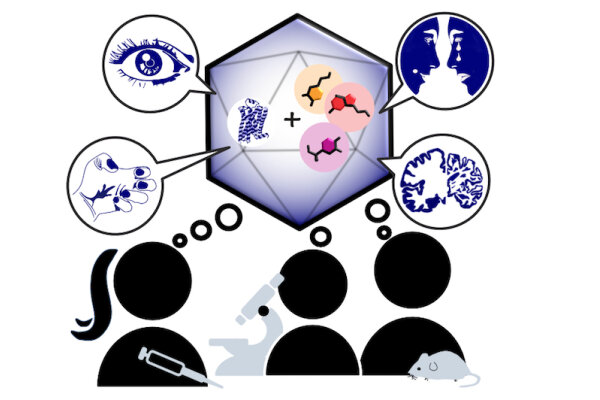
Monoaminergic transmitter systems originate in defined nuclei within the midbrain/hindbrain regions. The transmitter systems comprise of small numbers of neuronal clusters (i.e. 10,000 – 30,000 cells in rodents), which project into all brain regions. Some of these neurons generate up to over 200,000 synapses and release transmitter over large areas of the brain. Thus, these transmitters can globally modulate brain regions to adapt neuronal network function to changes of environmental cues associated with, for example, stress, fear, aversion, reward, or disease (Bear et al., 2015). In particular, neuromodulation of brain activity can lead to a switch between behavioral states such as sleep, attention or arousal (Lee and Dan, 2012; Noudoost and Moore, 2011; Shine, 2019). Thus, neuromodulation of neuronal networks can enhance or dampen information processing and therefore stabilize or destabilize learning processes and memory formation. Because of its importance in setting brain states, abnormal changes in the neuromodulatory capacity of a neuronal network (i.e. increased or decreased modulatory tone) either during development or in adulthood, may cause neuropsychiatric diseases, such as anxiety disorders, chronic depression and schizophrenia. Furthermore, deficits in neuromodulatory systems are currently emerging as risk factors for neurodegenerative disorders such as PD, AD and retinal disease (van Doze and Perez, 2012; Huang et al., 2017; Pardue and Allen, 2018). In addition, monoaminergic transmitters are essential for the regulation of movement and basal muscle tone. Therefore, malfunctions of monoaminergic transmitter systems are associated with movement disorders such as dystonia and PD (Ng et al., 2015). Despite this knowledge, therapy of affective brain disorders and monoamine-related dysfunction has not advanced for well over a decade. The same strategies are still used that were developed in the 1950s (dopamine receptor modulation in schizophrenia), 1980s (catecholamine reuptake inhibition for affective disorders such as depression and anxiety) and 1990s (serotonin reuptake inhibition for affective disorders). We believe that this is because the precise action of monoaminergic GPCRs in modulating brain function is neither fully understood, nor has it been adequately characterized due to the lack of tools to monitor their spatiotemporal activity.
In the last decade pharmacogenetics and optogenetic techniques have revolutionized our ability to control and understand the function of neuronal networks, cell-types and signaling cascades in relation to behavior (Rost et al., 2017; Song and Knöpfel, 2016; Wiegert et al., 2017). Optogenetics use light-activated proteins derived from animals, plants, fungi or bacteria to control ion fluxes, protein-interactions or intracellular signals in cells by light. Pharmacogenetics use chemical compounds, which can be tethered to the cellular target of choice and can also be light-switchable to control ion channels, protein interactions, intracellular signals and also membrane lipids. In combination with cell-type specific promotors using Cre-recombinase techniques, newly developed virus approaches and application of these methods in various animal models, these tools have shed light onto the function of single neurons and neuronal populations for animal behaviors and disease states. While optogenetics and pharmacogenetics allow the precise control of cellular events in a cell-type specific manner, genetically encoded sensors are used to visualize voltage changes, ion fluxes and signaling events. These new tools (sensors and switches) now allow us to control, visualize and analyze monoaminergic transmittersystems in neuronal circuits in a new dimension, to gain an understanding of the functional interaction of dopamine, serotonin and noradrenaline, their receptors and their activated intracellular signals and how these signals change during diseases related to monoaminergic transmitters.
Copyright © monndi 2025
Last update: Apr 22, 2025


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}