





Dr. Markus Piotrowski
Ars legendi Faculty Award
 Congratulations to Markus Piotrowski from our group for winning the Ars legendi Faculty Award for Excellent Teaching in Biology 2019! Stifterverband-Webseite | press release RUB
Congratulations to Markus Piotrowski from our group for winning the Ars legendi Faculty Award for Excellent Teaching in Biology 2019! Stifterverband-Webseite | press release RUB
Biosynthesis of the metal chelator molecule nicotianamine
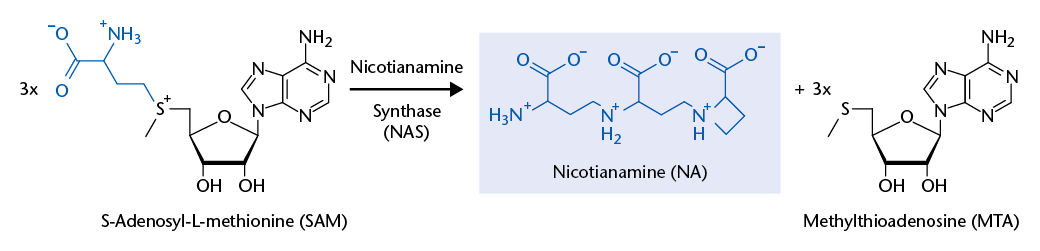
Background and preliminary work: The Krämer group has discovered that a plant membrane transport protein, Arabidopsis thaliana Zinc-Induced Facilitator 1 (ZIF1), mediates the vacuolar sequestration of nicotianamine (NA). NA is a non-proteinogenic amino acid that acts as a high-affinity metal chelator molecule in metal homeostasis of plants and filamentous fungi. Furthermore, NA has promising anti-hypertensive pharmacological activity. Roots of Arabidopsis thaliana overexpressing AtZIF1 accumulate ~30-fold enhanced NA concentrations. Nicotianamine synthases (NAS) of plants and filamentous fungi convert three S-adenosylmethionine (SAM) molecules to one NA molecule. NAS catalyses several consecutive reactions in a single active site, as concluded based on crystal structures of an NAS-like archaeal thermoNAS. Its reaction product, thermoNA, contains a glutamate instead of the terminal SAM-derived unique azetidine-2-carboxylate moiety that is characteristic of NA.
NAS proteins were first identified in barley and other plant species, and only later in filamentous fungi. More recently, NAS-like genes were found to be part of a Pseudomonas aeruginosa operon essential for survival on human airway mucus secretions, and to act in the synthesis of an NA-related compound of Staphylococcus aureus that utilizes only a single SAM molecule. Interestingly, no proteins acting in the catabolism of NA are known. A possible NA breakdown product – the toxic proline analogue and allelopathic chemical azetidine-2-carboxylate – was detected in only a few plants to date.
CN Hydrolases of Plants
![]() Our group is interested in the analysis of the so-called C-N hydrolases. These enzymes are involved in the catalysis of different types of C-N bonds like amides, imides, and nitriles (EC. 3.5.x.x: hydrolases, cleaving C-N bonds but not peptide bonds). C-N hydrolases are phylogenetically related and have to be distinguished from other enzymes which do also hydrolyse C-N bonds (e.g. peptidases), but share no common ancestor to the C-N hydrolases.
Our group is interested in the analysis of the so-called C-N hydrolases. These enzymes are involved in the catalysis of different types of C-N bonds like amides, imides, and nitriles (EC. 3.5.x.x: hydrolases, cleaving C-N bonds but not peptide bonds). C-N hydrolases are phylogenetically related and have to be distinguished from other enzymes which do also hydrolyse C-N bonds (e.g. peptidases), but share no common ancestor to the C-N hydrolases.
![]() Even the small genome of the model plant Arabidopsis thaliana (sometimes called “the green Drosophila”) possesses 9 genes for different C-N hydrolases. At the moment our focus is directed towards the nitrilases (4 different genes in Arabidopsis) and the involvement of C-N hydrolases in the biosynthesis of polyamines.
Even the small genome of the model plant Arabidopsis thaliana (sometimes called “the green Drosophila”) possesses 9 genes for different C-N hydrolases. At the moment our focus is directed towards the nitrilases (4 different genes in Arabidopsis) and the involvement of C-N hydrolases in the biosynthesis of polyamines.
1. Nitrilases
![]() Nitrilases (EC 3.5.5.x) catalyze the hydrolytic cleavage of nitriles (R-CN, also known as cyano-compounds) to their corresponding carboxylic acids. The four nitrilases of A. thaliana can be divided into two groups, according to their homology and their biochemical characteristics: NIT1-NIT3 (NIT1-group) and NIT4.
Nitrilases (EC 3.5.5.x) catalyze the hydrolytic cleavage of nitriles (R-CN, also known as cyano-compounds) to their corresponding carboxylic acids. The four nitrilases of A. thaliana can be divided into two groups, according to their homology and their biochemical characteristics: NIT1-NIT3 (NIT1-group) and NIT4.
1.1. Evolution of nitrilases in glucosinolate-forming plants
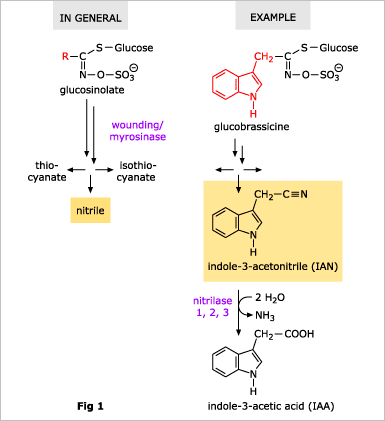
![]() In addition to A. thaliana, NIT1-group nitrilases are only known from two Brassica species. These three plant species belong to the family of the Brassicaceae. This family is also well known for their high content of glucosinolates. If plants are wounded, glucosinolates are released from their cellular stores and hydrolyzed by thio-glucosidases (syn. myrosinases). The final products of these reactions are, depending on special proteins present in the plant, isothiocyanates, thiocyanates, or nitriles (Fig. 1). A. thaliana contains at least 20 different glucosinolates, therefore 20 different nitriles can be formed. Interestingly, the NIT1-group nitrilases have a broad substrate-specificity and may be able to use this nitriles as substrates (Fig. 1).
In addition to A. thaliana, NIT1-group nitrilases are only known from two Brassica species. These three plant species belong to the family of the Brassicaceae. This family is also well known for their high content of glucosinolates. If plants are wounded, glucosinolates are released from their cellular stores and hydrolyzed by thio-glucosidases (syn. myrosinases). The final products of these reactions are, depending on special proteins present in the plant, isothiocyanates, thiocyanates, or nitriles (Fig. 1). A. thaliana contains at least 20 different glucosinolates, therefore 20 different nitriles can be formed. Interestingly, the NIT1-group nitrilases have a broad substrate-specificity and may be able to use this nitriles as substrates (Fig. 1).
![]() Due to the evolution of the glucosinolate-biosynthesis enzymes, different species may form a different spectrum of glucosinolates and, accordingly, different glucosinolate-derived nitriles. We want to investigate, if, to what extent, and how the nitrilases of glucosinolate-forming plants have evolved to match their proposed new substrates, the glucosinolate-derived nitriles.
Due to the evolution of the glucosinolate-biosynthesis enzymes, different species may form a different spectrum of glucosinolates and, accordingly, different glucosinolate-derived nitriles. We want to investigate, if, to what extent, and how the nitrilases of glucosinolate-forming plants have evolved to match their proposed new substrates, the glucosinolate-derived nitriles.
![]() An important aspect of the glucosinlate/nitrilase-system is that a certain glucosinolate, glucobrassicin (indolyl-3-methyl-glucosinolate), can release indole-3-acetonitrile (IAN), which, in turn, can be hydrolyzed to the plant hormone indole-3-acetic acid (IAA) by the nitrilases. This means that in plants which contain IAN (or glucobrassicin), nitrilases may have an important role in the homeostasis of IAA.
An important aspect of the glucosinlate/nitrilase-system is that a certain glucosinolate, glucobrassicin (indolyl-3-methyl-glucosinolate), can release indole-3-acetonitrile (IAN), which, in turn, can be hydrolyzed to the plant hormone indole-3-acetic acid (IAA) by the nitrilases. This means that in plants which contain IAN (or glucobrassicin), nitrilases may have an important role in the homeostasis of IAA.
1.2. Involvement of nitrilases in cyanide detoxification in plants
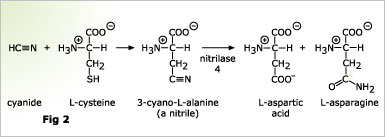
![]() In contrast to the NIT1-group nitrilases which seem to be restricted to the order of Capparales, NIT4 homologs seem to be widely distributed among higher plants. NIT4 of Arabidopsis and tobacco have a high substrate specificity for 3-cyano-L-alanine (cyanoalanine), an intermediate product of cyanide detoxification in higher plants. Cyanoalanine is formed by the coupling of cyanide to the amino acid L-cysteine by the enzyme cyanoalanine synthase (Fig. 2). Many years ago an enzyme was described, cyanoalanine hydrolase, which further converts cyanoalanine to asparagine. We wonder if cyanoalanine hydrolase (the sequence of which is unknown) is identical to NIT4.
In contrast to the NIT1-group nitrilases which seem to be restricted to the order of Capparales, NIT4 homologs seem to be widely distributed among higher plants. NIT4 of Arabidopsis and tobacco have a high substrate specificity for 3-cyano-L-alanine (cyanoalanine), an intermediate product of cyanide detoxification in higher plants. Cyanoalanine is formed by the coupling of cyanide to the amino acid L-cysteine by the enzyme cyanoalanine synthase (Fig. 2). Many years ago an enzyme was described, cyanoalanine hydrolase, which further converts cyanoalanine to asparagine. We wonder if cyanoalanine hydrolase (the sequence of which is unknown) is identical to NIT4.
2. Biosynthesis of putrescine in plants
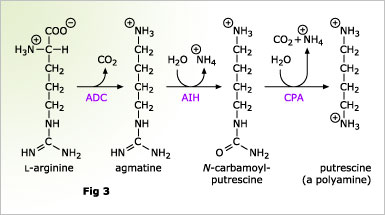
![]() Putrescine is a so-called polyamine, which is a group of certain small oligoamines found in every living organism. For plants important functions like control of cell division and response to salt stress were assigned to this molecules. In plants two different routes of polyamine biosynthesis have been described, but, at least in Arabidopsis, one of this pathways, the ornithine decarboxylase pathway, is missing. The second pathway, called the arginine decarboxylase pathway, starts with the decarboxylation of the amino acid L-arginine. The resulting agmatine is subsequently hydrolyzed to N-carbamoylputrescine by agmatine iminohydrolase (AIH) and further to putrescine by N-carbamoylputrescine amidohydrolase (CPA)(Fig. 3).
Putrescine is a so-called polyamine, which is a group of certain small oligoamines found in every living organism. For plants important functions like control of cell division and response to salt stress were assigned to this molecules. In plants two different routes of polyamine biosynthesis have been described, but, at least in Arabidopsis, one of this pathways, the ornithine decarboxylase pathway, is missing. The second pathway, called the arginine decarboxylase pathway, starts with the decarboxylation of the amino acid L-arginine. The resulting agmatine is subsequently hydrolyzed to N-carbamoylputrescine by agmatine iminohydrolase (AIH) and further to putrescine by N-carbamoylputrescine amidohydrolase (CPA)(Fig. 3).
![]() The activity of the latter two enzymes in plants is known for a very long time, but only recently we have cloned and characterized both enzymes from Arabidopsis. Currently we are studying these enzymes from other organisms.
The activity of the latter two enzymes in plants is known for a very long time, but only recently we have cloned and characterized both enzymes from Arabidopsis. Currently we are studying these enzymes from other organisms.
Further information
For further informations please visit my "C-N-Hydrolase Group"-Homepage.
![]()


